 网硕互联帮助中心
网硕互联帮助中心《智能的理论》全书转至目录
不同AGI的研究路线对比简化版:《AGI(具身智能)路线对比》,欢迎各位参与讨论、批评或建议。
一.视觉通路概述(Goldstein和Brockmole(著),张明(译),2018)
视觉感知的基本过程如下:首先,视觉的起点源自于现实世界中的物体。光线从物体表面反射,进入眼睛,而眼睛作为视觉或光线的感受器,接收到这些光线。光线在进入眼球后,在视网膜上形成物体的图像。在视网膜层面,视觉感受器将光信号转换为电信号,并传递至大脑。随后,在大脑中,特征察觉器将这些电信号解析为各种特征,例如颜色特征、大小特征、线条特征、轮廓特征等,这一过程可视为特征的分解。最终,大脑将这些特征重新整合,形成对物体的类别知觉等,这一步骤是特征的重新整合过程。此外,注意力和记忆在视觉感知中扮演着至关重要的角色。
感觉和知觉是经常被用于,有必要需要区分一下它们。感觉是个体通过感官(如眼、耳、鼻、舌、皮肤等)直接接收外界刺激后,在大脑中产生的对事物个别属性的反映,发生在感觉系统的初期加工阶段。例如:看到苹果的红色(视觉属性)、摸到苹果的光滑(触觉属性)、闻到苹果的清香(嗅觉属性)。知觉是个体在感觉的基础上,对事物的整体属性进行综合加工和解释的过程,它是一种高水平的复杂加工,涉及与脑活动相关的解释与记忆。例如,当我们同时接收到苹果的红色、光滑、清香等多个感觉信息时,会将这些信息整合起来,并与记忆等高级加工水平综合起来,以此判断 “这是一个苹果”,这种对事物整体的认知就是知觉。
感觉与知觉并不泾渭分明,比如看到一个点,加上这个观察并没有涉及高水平的加工,此时发生的就是感觉,但如果这个点涉及到解释等,如“这个点是一张白纸中间的一个洞”,此时就变成了知觉。
二.眼睛(Goldstein和Brockmole(著),张明(译),2018)
眼睛是动物视觉系统的接收装置,位于视觉通路的首端。它由视网膜、角膜、晶状体、瞳孔等多个部分构成,能够处理波长范围在400至700纳米的电磁波,即我们所见的可见光。
1.视网膜
光线在进入眼睛后,会在视网膜上呈现出一个图像。 随后,视网膜上的视觉感受器将这一图像转换为电信号。视网膜成像的原理基于小孔成像,因此所形成的图像是倒立的。在此过程中,外界物体被称为远刺激,而其在视网膜上的成像则被称为近刺激,如图。
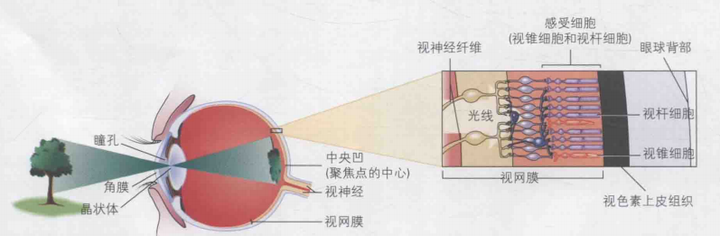
图1
2.角膜、晶状体和瞳孔
人的眼睛具有调节的功能,能够看清不同距离的物体,这一功能主要由角膜、晶状体和瞳孔三个组织协同完成。光线进入眼睛后,依次经过角膜、瞳孔和晶状体。角膜和晶状体的主要作用是聚焦光线,它们能够将物体上的某一点发出的多条光线聚焦在视网膜的对应点上。其工作原理类似于凸透镜,通过对光线的折射实现聚焦,在眼科学中,这种折射被称为“屈光”。其中,角膜提供了约80%的聚焦能力,剩余的20%则由晶状体承担。角膜与晶状体在功能方面存在差异:角膜的聚焦能力是固定的,而晶状体则能通过睫状体的放松和收缩来调节聚焦能力。如图2所示,当观察远处物体时,睫状体放松,晶状体变薄,屈光力减弱,使远处物体的平行光线焦点落在视网膜上(超过6米远的物体的反射光线基本平行),如图2(a)。此时,由于屈光力度不足,近处物体的焦点会落在视网膜后方,导致近处物体看起来模糊。相反,若要观察近处物体,睫状体需收缩,使晶状体变厚,屈光力增强,从而使近处物体的焦点落在视网膜上,如图2(b)。此时,远处物体的焦点则会落在视网膜前方,导致远处物体看起来模糊。近视的常见原因在于角膜和晶状体对光线的过度弯曲,使远处光线聚焦在视网膜之前,因此近视患者难以看清远处物体;而远视患者则能看清远处物体,但却难以看清近处物体,因为光线更容易聚焦在视网膜后方。晶状体的调节过程无需意识参与,使得物体聚焦过程极为高效。瞳孔则负责调节进入眼球的光线量,当观看远处物体时,瞳孔会扩大以允许更多光线进入;观看近处物体时,瞳孔则收缩。
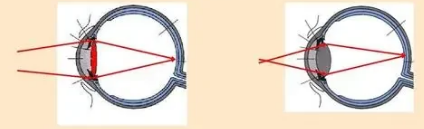
图2(a) 图2(b)
3.视觉感受器
视觉感受器主要由视锥细胞和视杆细胞构成,它们分布在视网膜上,其核心功能是将光信号转换为电信号。视网膜可划分为两个主要区域:其一是中央凹,位于视网膜中心位置,但它仅占据视网膜的极小部分;其余则为外周视网膜,具体分布情况如图3所示。
眼球上大约有600万个视锥细胞和1.2亿个视杆细胞。其中,视锥细胞主要集中于极小范围的中央凹区域,并在中央凹处具有密度较高(约5万个),而在外周视网膜上密度较低;相反,视杆细胞则主要分布在大范围的外周视网膜上,如图3(b)所示。
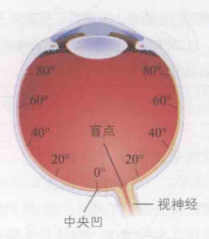
图3(a)
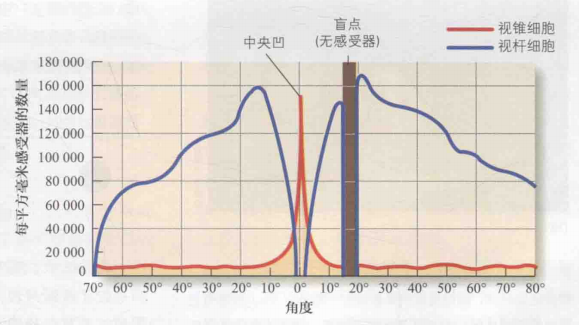
图3(b)
电信号生成后,依次传递至双极细胞和神经节细胞,随后神经节细胞通过视神经纤维(即神经节细胞的轴突,长度较长)将信号传送至大脑。值得注意的是,如图4,双极细胞和神经节细胞覆盖在视觉感受器的外面,此外还分布有水平细胞和无足细胞,光线正是透过这四种神经细胞抵达视觉传感器。另外,视神经纤维从外向内穿过视网膜,形成了一个盲点。然而,这一盲点通过我们大脑的高级神经处理过程得以“脑补”,因此我们感知不到这个盲点的存在。
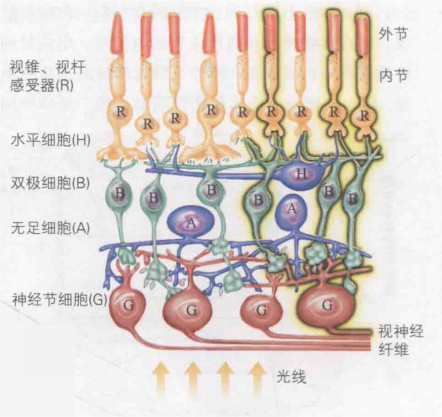
图4
视觉感受器通过神经节细胞的视神经纤维将信号传递至大脑。由于一只眼睛拥有1.26亿个感受器(包括1.2亿个视杆细胞和600万个视锥细胞),而神经节细胞仅有100万个,因此平均每个神经节细胞需接收来自126个感受器细胞的信号。这种一个神经节细胞接收多个感受器信号的现象称为聚合。两种视觉感受器的聚合情况是不同的。视锥细胞的聚合程度要远低于视杆细胞(相较于视锥细胞,一个神经节细胞会聚合更多的视杆细胞)。特别是在中央凹区域,一个神经节细胞仅接收一个视锥细胞传递的信号,无需聚合。聚合程度的差异导致两种感受器在功能上存在两点主要差异:(a)视杆细胞比视锥细胞更为敏感;(b)视锥细胞对视觉信息的处理比视杆细胞更为细致。
(a)视杆细胞比视锥细胞更灵敏
视杆细胞的聚合程度高于视锥细胞,因此外周视觉(富含高密度视杆细胞)比中央凹更容易察觉微弱光线。天文学家和业余占星爱好者熟知的一种现象可以佐证这一点:若直接注视亮度微弱的星星,它们往往难以被察觉(因星星图像落在仅分布视锥细胞的中央凹上);然而,若不直视,让星星偏离眼睛注视点,这些亮度微弱星星便能被觉察(因星星图像落在外周视网膜的视杆细胞上)。
产生这种这种现象的原因在于视杆细胞发生的聚合多于视锥细胞。可以通过下图5作进一步说明:每一个光子射入一个视锥细胞,并且每个视锥细胞联结一个神经节细胞。此时,若光子强度不足(如2个强度),则不足以引起神经节细胞的反应;而多个光子分别刺激多个视杆细胞,这些视杆细胞共同连接一个神经节细胞,每个光子的强度叠加起来(如10个强度),便足以引发神经节细胞的反应。因此,相比视锥细胞,视杆细胞在微弱光线条件下更容易产生反应。
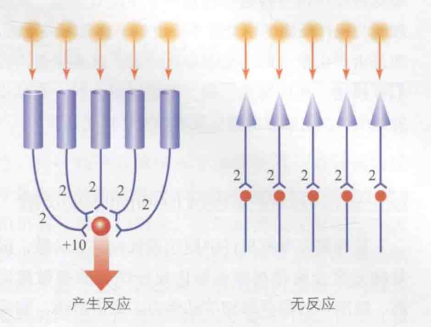
图5
(b)视锥细胞比视杆细胞有更高的视敏度
缺乏聚合作用使得视锥细胞的视敏度相较于视杆细胞更高。视敏度是指观察视觉细节的能力,简而言之,能够清晰辨认视力表上的微小字母,即代表视敏度高。显而易见,我们视网膜的中央凹的视敏度最为出色,因为只有当物体成像于中央凹时,我们才能清晰地看到物体,而物体若落在周边视觉区域,则会显得模糊不清。这主要归因于中央凹区域的视锥细胞仅与单一神经节细胞相连。具体的原理可通过下图6作进一步说明。
图(a)展示了视杆细胞的神经环路。当呈现两个相邻的光点刺激时(左图),视杆细胞会发出信号,引起神经节细胞放电。 若将两个光点分开(右图),此时两个分离的视杆细胞发射的信号仍会传递给同一个神经节细胞,并导致其放电。然而,大脑在接收到该神经节细胞的信号时,无法区分光点的照射状态(即左图还是右图)。
图(b)展示了视锥细胞的神经环路结构,其中每个突触均与相应的神经节细胞相连。当出现两个相邻的光点刺激时(如左图所示),相应的两个相邻神经节细胞会放电。而当将两个光点分开时(如右图所示),对应的两个分离的神经节细胞则会放电。由此可见,不同的光刺激模式会导致不同的神经节放电反应,从而使大脑接收到不同的信号,并据此进行区分。
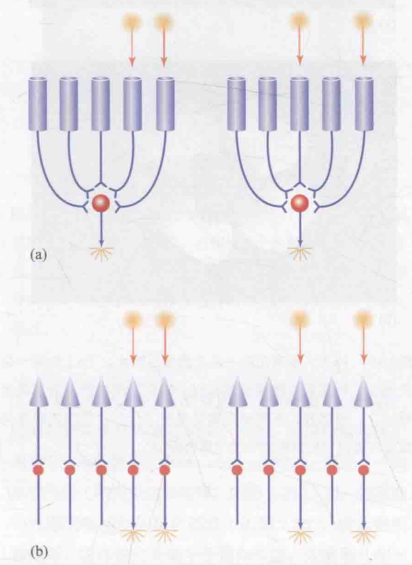
图6
4.视神经纤维
(1)感受野
视觉信号(电信号)在离开视网膜后,通过视神经纤维与大脑相连,如图7。这些神经纤维连接着视网膜上的特定区域,当该区域出现小光点时,相应的神经纤维就会产生放电反应。视网膜上的这一区域,被称为该神经纤维对应的感受野。这些感受野有的部分重叠,有的甚至完全重叠(即一些不同的神经节细胞覆盖的感受野位置重叠)。人类单只眼睛大约拥有一百万根视觉神经纤维,这些数量庞大的视觉神经纤维的感受野共同构成了整个视网膜图像,从而呈现出完整的视野。感受野这一概念由美国生理学家H. Keffer Hartline发现并命名,他在用光照射青蛙视网膜的极小区域时,观察到相应的视觉神经纤维被激活。视觉信号从这些视神经纤维出发,继续向大脑传递,部分大脑皮质的神经元汇聚了多个视觉神经纤维的信号,这意味着这些更高级的大脑神经元的感受野覆盖了其底层神经元的感受野,因而具有更大的感受野,并整合了更多的视觉信息。
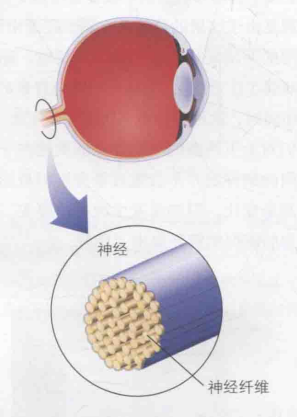
图7
我们可以通过一个物流(快递)系统来类比感受野。假设每个物流网点对应一个街道,那么可以认为一个最小层次的感受野相当于一个街道,而神经节细胞则相当于物流网点。一个城市站点管理该城市内的所有网点,因此一个较大的感受野可以类比为整个城市,一个较深层的神经元则相当于城市的物流站点。依此类推,到省级层面亦是如此。当所有街道、城市及省份拼接在一起时,便形成了完整的地图,这相当于我们的整个视野。
一般来说,神经元的感受野都是固定的。但是也存在一些例外情况。Womelsdorf及其同事(Womelsdorf,Anton-Erxleben,Pieper和Treue,2006)在对猴子的颞叶神经元的记录过程中证明,注意可以改变神经元感受野的定位。图8展示了一个神经元感受野所在的位置,当时,这只猴子正注视着屏幕左上方的白点,同时又注意着箭头所指的菱形位置。图8最示了当这只猴子把注意转移到箭头所指的圆圈位置时,感受野的位置的变化。在这个例子中,黄色代表了某视觉神经元接收刺激时产生最大反应量的视网膜区域。
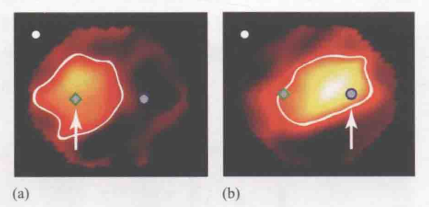
图8
(2)放电特征
后来的研究者们记录了猫的视神经纤维信号,并发现了感受野的“中心-周边结构”(Barlow,Fitzhigh和Kuffler,1957;Hubel和Wiesel,1965)。使用下图9可以说明这一结构,在图(a)中的感受野中,当光点照射中心区域时,放电活动增强,因此该中心区域被称为感受野的兴奋区。相反,刺激周边区域会导致放电率下降,故此区域被称为感受野的抑制区;而在图(b)的感受野中,情况则截然不同,中心区域受到刺激时表现为抑制,周边区域受到刺激时则表现为兴奋。
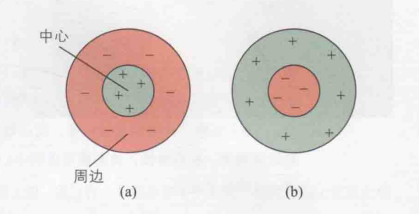
图9
一个感受野可以被划分为兴奋区和抑制区,信号刺激兴奋区会激活神经元反应,而信号刺激抑制区则会抑制反应。当兴奋区的受刺激面积增加时,反应强度也随之增强;相反,抑制区的受刺激面积增大,反应强度则会减弱。如图10所示,该感受野呈现出中心兴奋和周边抑制的特点,其中黄色部分代表受到光刺激的区域。每个感受野下方均标注了神经元对刺激的反应情况。如图(a)所示,当仅有一个小光点刺激时,产生的反应较小;图(b)展示了当整个兴奋区域受到刺激时,反应达到最强;如图(c)所示,随着光点面积继续增大,直至部分周边抑制区受到刺激,反应开始减弱,此图旨在说明中心与周边的拮抗作用;如图(d)所示,当光照覆盖全部周边抑制区时,反应进一步降低。这一现象可以通过侧抑制(9-7:轮廓)的原理来解释。
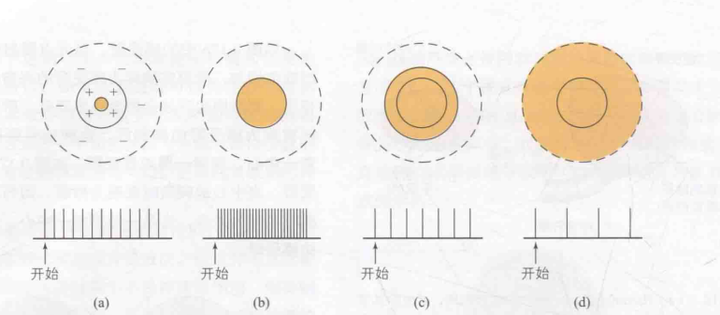
图10
三.外侧膝状体(注:来自《视觉系统的全貌》 https://zhuanlan.zhihu.com/p/123892438)
1.整体通路
视觉信号沿着视神经纤维传输后,首先会到达视交叉。视交叉就像是一个物流分选站,它把左眼的右侧视野和右眼的右侧视野打包在一起,并传入左侧大脑;而左眼的左侧视野和右眼的左侧视野,被打包在一起,传入右侧大脑。然后视觉信号会达到外侧膝状体(需要记住,左视野信息到达右脑的外侧膝状体,右视野信息到达左脑的外侧膝状体)。最后,视觉信号经外侧膝状体到达大脑皮层枕叶。外侧膝状体接收了90%的离开眼睛的视神经纤维,另外10%到达上丘,上丘是和眼动相关的重要脑区,参与由视觉、听觉等外界因素引起的眼动反射活动。
相比视神经纤维,外侧膝状体神经元的感受野只发生了微小的变化。另外,它本身就是一个包含了上百万神经元的复杂结构。因此在外侧膝状体中,总应该有一些事情发生。大脑皮质也会向外侧膝状体传递反馈信号,且要多于来自视网膜的信号,如图11。因此对外侧膝状体有一种猜想:从大脑传递回外侧膝状体的信息可能影响了哪些信息会被上传到大脑。
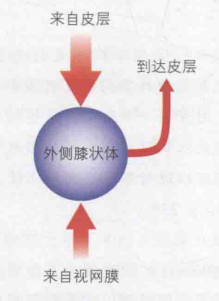
图11
2.结构
与视神经纤维非常类似,外侧膝状体的感受野也是中心-周边结构,中心是兴奋性的而周边是抑制性的,或相反。另外,外侧膝状体具有分层结构,每一层的细胞都有不同的作用。人的外侧膝状体分为六层,从下到上分别命名为第1、2、3、4、5、6层,且每层都仅表征对侧视野(左脑外侧膝状体的每一层组织与(左眼及右眼的)右视野对应;右脑外侧膝状体的每一层组织与左视野对应。例如,如图左边的外侧膝状体接收的是右侧视野的蛋糕)。当然这种对应是拓扑学上的,即在视网膜图像上相邻的点,在外侧膝状体里形成的图像上依然是相邻的,即只考虑物体间的相对相邻关系而不考虑它们的形状和大小(位置也存在一定的形变,仅存在相邻关系)。最后,外侧膝状体的神奇之处在于:同侧眼睛发来的信号,由2、3、5层细胞接收;对侧眼睛发来的信号,由1、4、6层细胞接收,如图12。
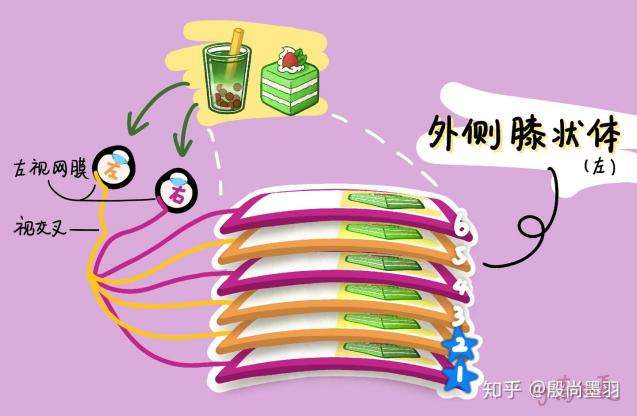
图12
外侧膝状体的拓扑图有一个特点,越靠近中央凹的位置,物体所占拓扑图的面积越大。其中的一个原因是,外周视野通常几个视觉感受器对应一个神经节细胞,因此图像相应的会被缩小;相反,中央凹的一个视觉感受器对应一个神经节细胞。当然,中央凹视觉是最需要被注意的信息,因此理所当然的需要获取更多的资源(拓扑图面积)。
3.分拆与重组
外侧膝状体有6层,每层都是与视野对应。那么这6层信息是如何整合起来传给大脑皮层的呢?
事实上,每层组织与视网膜的拓扑结构都是相同的。以下图13作为说明,把外侧膝状体的每一层,都当作一张世界地图,且每个地图对应位置的图像内容都相同,比如,第一张地图的某个位置是新西兰,其他5张地图的对应位置也是新西兰(来自左眼的信号用红色表示;来自右眼的信号用蓝色表示)。
外侧膝状体在上传给视觉皮质之前,每层会按照位置拆分为不同的“碎片”,然后这6层对应位置的碎片会被打包在一起,然后从这个“包裹”里抽取一个碎片,作为包裹里6个碎片的代表。例如,在每层地图上将新西兰(都是对应位置)作为一个碎片取出,得到6个新西兰碎片,然后再抽取一块新西兰碎片。当抽取完整个视野的碎片时,便重新组合得到整个视野。然后这个视野信息被一次性交给了初级视觉皮层。
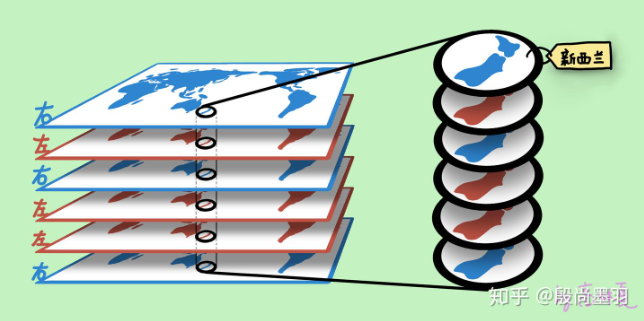
图13
4.大细胞层和小细胞层
外侧膝状体内部分为6层,其中第1、2层和3、4、5、6层分属两类——大细胞层和小细胞层。所谓大细胞,指的是这类细胞的感受野尺寸比较大。同理,小细胞的感受野比较小。大细胞和小细胞分别对不同类型的视觉信号敏感,大细胞对运动的方向、速度信息很敏感,但是对物体的精细结构不敏感(检测运动方向信息和速度信息);小细胞喜欢对物体的精细结构敏感,但对运动信息不敏感(检测长条物的方向信息和形状信息)。获得这些视觉信息后,大、小细胞就将这些视觉信号分为两类,各自打包向皮层传输,从而实现了“双通道并行”的模式。
四.初级视觉皮质(V1区)
初级视觉皮质,即V1区,位于枕叶,因其横断面呈条纹状,故又被称为纹状皮层。它是大脑皮质中最早接收来自眼球和外侧膝状体视觉信息的区域。
1.特征察觉器
特征察觉器是一种对特定特征(如颜色、线条方向、形状)产生反应的细胞。神经节细胞的感受野能够对中心或周围区域做出响应,因此它们属于特征察觉器。当信号传入外侧膝状体后,该区域的神经元同样拥有与神经节细胞相似的感受野,故而这些细胞也被视为特征察觉器。
神经科学家Hubel和Wiesel(1959)在初级视觉皮质中发现了皮层简单细胞。这类细胞对其感受野内出现的特定方向的线条产生反应,不同的皮层简单细胞对不同角度的线条有特定的反应,例如存在对45度角方向的线条反应的神经元或对垂直方向的线条反应的神经元。进一步研究表明,当线条方向与该细胞对应的方向一致时,放电率最大;随着线条方向逐渐偏离该方向,放电率会逐渐减弱。例如,检测垂直方向的特征察觉器对垂直线条(90°)的反应最强,而对80°线条的反应则相对较弱,依此类推,直至0度时处于静息状态。神经元的激活状态并非仅有激活和静息两种,其激活程度是一个连续变化的值,且与刺激与特征察觉器的匹配程度呈正相关关系。
除此之外,该研究团队在初级视觉皮质及其邻近的视觉区域发现了皮层复杂细胞。这类细胞能够对特定朝向及向特定方向运动的线条产生反应。具体而言,当一个特定朝向的线条在其感受野中沿某个特定方向运动时,该细胞会被激活。例如,对-45度朝向且正朝45度角方向运动的线条,这类细胞会有显著反应。这类细胞被称为皮层复杂细胞。此外,还存在一种端点细胞,它们对特定长度、特定朝向和特定运动方向的线条(如45度切斜且朝-45度角方向运动的短线条)产生反应,或者对角形图案(即端点)产生反应。如图14所示。
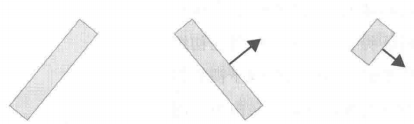
图14
2.定位柱与方向柱
在初级视觉皮质中,这些特征察觉器是如何组织的呢?在Hubel和Wiesel(1965)进行的研究中,研究人员发现了定位柱。定位柱是一块大约为1毫米大小的皮质区域(垂直于初级视觉皮质表面的一小块),在初级视觉皮质中平铺着大量的定位柱。在同一块定位柱内,其内部的所有细胞(即特征察觉器)的感受野大致相同。此外,他们还发现定位柱中存在许多方向柱(方向柱又是定位柱内的一小块区域)。但需要注意的是,同一块定位柱中的所有方向柱的感受野相同,但每个方向柱检测的线条方向不同,即仅对同一位置不同方向的线条产生反应。同时,相邻方向柱对应的最佳方向相似。例如,某个方向柱的最佳方向是90°,那么其相邻方向柱的最佳方向可能是89°…。如图15所示,蓝色代表定位柱,它是一小块纹状皮层,尺寸约为1毫米。其中的黄色纹状皮层即为方向柱。当一个定位柱包含所有方向的方向柱时,该定位柱能够对所有方向的线条产生反应,被称为超柱。
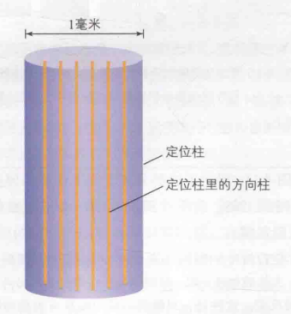
图15
3.视网膜脑图
视网膜在大脑皮质上的位置分布图被称为视网膜脑图,它展示了某块皮质与视网膜上特定位置的对应关系。如图16所示,当一个人注视一棵树时,树上的A、B、C、D点分别对他视网膜上的A、B、C、D四点产生了刺激。进一步观察大脑皮层,可以发现视网膜图像中的A点引发了大脑皮层中A点神经元的放电反应,同理,视网膜中的B点引发了B点神经元的放电,以此类推。
此外,视网膜上不同位置所对应的皮质面积存在差异。具体而言,中央凹区域的视网膜点对应的皮质面积较大,而外周区域的则相对较小。根据脑电测量的数据,尽管中央凹仅占视网膜总面积的0.01%,但其对应的视网膜脑图却占据了8%至10%。例如,图16中的C点和B点靠近中央凹,因此它们所占的皮质面积较大;而A点和D点远离中央凹,所占皮质面积较小。这种差异的产生原因在于,中央凹需要处理更多的视觉细节,因而需要更多的皮质资源来支持。
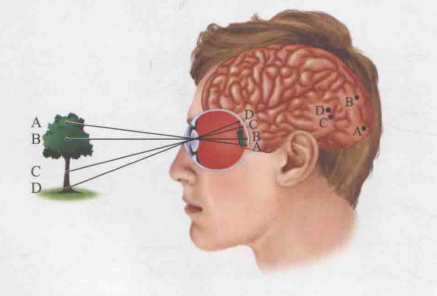
图16
4.平铺效应
初级视觉皮质是如何对整个视野进行反应的呢?初级视觉皮质内含有众多定位柱,每个定位柱分别对应视野中的不同位置。这些定位柱相互协作,共同作用,从而实现对整个视野的全面覆盖,这一现象被称为平铺效应。对于长条形物体,能够激活相邻定位柱中相同方向的方向柱,而这些方向柱必须被有效组织,才能形成对该物体的完整知觉。
如图17所示,黄色圆圈象征着各定位柱的感受野,当这些感受野相互叠加时,便能完整覆盖整个场景。如下图18所见,A区域、B区域和C区域分别对应树干的三个特定部位及其相应的三个定位柱。每个定位柱内含众多对特定方向敏感的方向柱,然而鉴于树干主要由垂直线条构成,仅对垂直线条敏感的方向柱会呈现出最强的放电反应,图19中以黄色线条标示。正是这三个方向柱的协同作用,共同构建了树干的知觉模型。

图17

图18
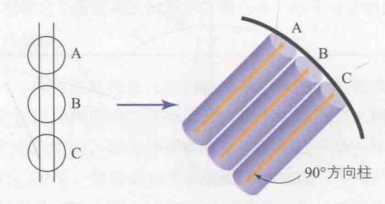
图19
五.腹侧通路与背侧通路
识别和定位是视觉的两大核心任务。为了确定识别和定位功能对应的脑区,Ungerleider和Mishkin(1982)对猴子进行了大脑切除术(如果丧失某项功能,即可确定被切除的脑区负责该功能),并设计了两项测试:客体识别任务和地标辨别任务。在客体识别任务中,猴子需要识别两个不同形状的物体,一个是目标物体(如长方体),另一个是非目标物体(如三棱柱)。若猴子能识别目标物并移开目标物体,即可获得食物奖励,如图20(a)。在地标辨别任务中,猴子的两侧各有一个盖子,当猴子移开距离圆柱体最近的盖子时,便能获取食物,如图20(b)。从枕叶的初级视觉皮层出发,存在两条主要通路:一条通往颞叶,称为腹侧通路(位于大脑腹部);另一条通往顶叶,称为背侧通路(位于大脑背部)。实验结果显示,当猴子的部分颞叶被切除时,在客体识别任务中会遭遇困难,因此可以确定颞叶与客体识别密切相关,腹侧通路也因此被称为内容通路或“what”通路。同理,当顶叶被切除时,猴子在地标辨别任务中会遇到困难,因此可以确定顶叶与定位功能相关,背侧通路则被称为位置通路或“where”通路。

图20(a)

图20(b)
视觉是服务于行为的,为的是能更好的和世界互动。因此Milner和Goodale(1995)认为,背侧通路与动作的实施有关。位置通路除了有空间信息,还涉及与物体的交互。为了证实这个观点,他们设计了如下实验。DF是一位脑颞叶的脑损伤者,实验要求她将卡片与插槽对齐但不用插入。这是一个识别任务,因为颞叶受损导致她无法辨别线条,所以她的对齐位置非常分散。如图21(a)左侧所示,而右边是正常人的测试结果。另外一个任务是将卡片插入插槽,因为她无法对齐所以她理应也无法插入。但是她做到了,如图21(b)所示。对此的解释是,识别和动作是双分离,即一个功能的损失未必会导致另外一个功能的损失。即便腹侧通路受损(无法识别方向),也不影响插入任务。基于此,研究者认为背侧通路更应被称为动作通路或者方式通路。

图21
六.其他脑区
除了皮层简单细胞、皮层复杂细胞和端点细胞,这里还存在更为复杂的特征察觉器,这一发现归功于Gross等人(Gross,Rocha-Miranda和Bender,1972)。研究人员对麻醉后的猴子进行了实验,他们将各种图形投影到屏幕上,以测试猴子颞下皮层的神经元反应。起初,他们仅使用线条、正方形和圆形等简单图案进行测试,但发现神经元对这些图案并无明显反应。几天后,一名实验人员无意中将手影投射到屏幕上,结果颞下皮层的某个神经元突然剧烈放电。随后,他们尝试了多种不同图案,发现图案与手越相似,神经元的反应就越强烈。此外,颞下皮层中还包含一个梭状回脸孔区,专门负责对脸孔的反应。同时,还存在对建筑、动物、风景等复杂图案反应的高级神经元。
七.感觉编码(Goldstein和Brockmole(著),张明(译),2018)
1.编码方式
感觉的神经表征,即大脑的放电模式,被称为感觉编码。目前存在三种主要的感觉编码假设。第一种是特异性编码,指的是一个细胞仅会被一个特定的物体激活。例如,在图22(a)中,不同的细胞分别被不同的人激活:比尔对应4号细胞,玛丽对应9号细胞,拉斐尔对应6号细胞,而其他细胞则处于静息状态。这种高度特异化的细胞形象得就像当遇到关于祖母的概念时细胞会激活,因而也被称为“祖母细胞”。第二种是群体编码,它认为任何一个物体都会引发大量特征察觉器的反应,但不同物体引起的反应模式各不相同,从而实现不同物体的区分。如图22(b)所示,尽管三人都激发了多个细胞的反应,但他们所对应的反应模式各有差异。最后一种是稀疏编码,与前两种编码方式相比,这种编码仅激活部分特征察觉器(数量少于全部,但多于一个),其余的则保持静止,如图22(c)所示。由于群体编码和稀疏编码均涉及多个神经元的协同作用,它们也被称为分布式编码。

图22(a)

图22(b)

图22(c)
早期研究明确指出,大脑中的特定区域负责处理特定的知觉类型,例如面孔和手等。而功能性磁共振成像(fMRI)的扫描结果显示,不同类型的刺激确实能够引发大脑多个区域的兴奋,从而证实了分布式表征理论。梭状回面孔区作为面孔识别的一个关键模块,可以被面孔刺激激活,同时面孔刺激还能激活其他脑区,这一现象进一步支持了分布式表征的观点。对面孔的感知体验并非仅仅局限于识别目标为面孔(“那是一张脸”)。我们还能对面孔的多种相关元素产生反应:(a)情感层面(她在微笑,因此她可能很开心);(b)注视方向(她正看着我);(c)面孔各部分的活动方式(通过观察他嘴唇的动作,我能更好地理解他);(d)面孔的吸引力(他有一张英俊的面庞);(e)面孔的熟悉度(我记得在哪儿见过她)。这表明,面孔能够引发多种反应,而这些不同的反应与大脑中不同区域的兴奋性密切相关。
在分布式编码的框架下,部分神经元对刺激所代表的概念产生反应,而非针对图片的特定的视觉特征。例如,某个神经元会对Halle Berry产生反应,无论呈现的是她的照片、画像、她在蝙蝠侠电影中猫女扮相的照片,还是“Halle Berry”的文字。此外,还有一些神经元对Jennifer Aniston和Lisa Kudrow产生反应,原因在于她们均出演了电视剧《老友记》。
最后,我们的大脑中拥有多样化的特征察觉器。当我们观察一个物体时,既有负责检测圆形的特征察觉器可能被激活,也有识别运动的特征察觉器,还有感知大小的特征察觉器,以及辨别颜色的特征察觉器等等。这些特征察觉器相互协作,共同构成一次“大合唱”,并奠定对物体知觉的基础。
2.可塑性
这些特征察觉器并非与生俱来,而是具有后天可塑性。Blakemore和Cooper(1970)进行了一项实验,他们将一只小猫置于一个仅包含垂直线条的环境中。如果特征察觉器后天无法改变,那么无论小猫在该环境中生活多久,当它被移出该环境并面对各种朝向的线条时,理应像正常动物一样对各种朝向的线条都产生反应。然而,经过在该环境饲养5个月后,实验人员对小猫的皮层进行了测试,结果发现小猫最终仅保留了针对垂直方向线条产生反应的神经元。如图23所示。

图23




评论前必须登录!
注册